Szántó G. Tibor, Panyi György és munkatársaik Shaker-IR típusú feszültségfüggő káliumcsatornák S6 hélixében lévő pórusformáló aminosav oldalláncok állapotfüggő hozzáférhetőségét tanulmányozták, hogy betekintést nyerjenek a lassú inaktiváció molekuláris mechanizmusába. A patch-clamp technika feszültség-zár üzemmódjában, inside-out konfigurációban, különböző Shaker-IR ioncsatornákon végzett méréseik során a cisztein oldalláncokhoz kötődő, ún. tiol-reaktív reagensek (pl. MTSEA és MTSET) hozzáférhetőségét tanulmányozták az ioncsatorna különböző kapuzási állapotaiban, különös tekintettel az inaktivációs kinetikára. Eredményeik azt mutatják, hogy az ioncsatorna zárt és nyitott állapotában a pórustól „elzárt” cisztein-oldalláncok a lassú inaktiváció során hozzáférhetővé válnak az MTSEA számára, ami arra utal, hogy az ioncsatorna aktivációs és a szelektivitási szűrő által formált lassú inaktivációs kapu közötti kommunikációt (azaz csatolást) az S6 hélix régióban bekövetkező molekuláris átrendeződések közvetítik, melyet molekuláris dinamikai szimulációk is alátámasztanak. Az S6 régió molekuláris átrendeződése továbbá összhangban van az S6 lassú inaktiváció során bekövetkező, a hossztengelye körüli merev, rúdszerű forgásával is. Eredményeik biológiai jelentősége egyrészt, hogy a lassú inaktiváció kinetikai alapon szabályozza az aktiválható ioncsatornák azon hányadát, amelyek kritikusak az ingerelhető sejtek számára, melyhez fontos a lassú inaktivációt vezérlő kapuzási folyamatok és konformációs változások minél jobb megértése. Másrészt, a KV ioncsatornák kapuzási átmeneteinek megértése elősegítheti az ioncsatornákhoz állapotfüggő módon kötődő, hatékonyabb gyógyszermolekulák, például antiaritmiás gyógyszerek tervezését is. Eredményeik a Journal of General Physiology (IF: 3.8) nemzetközi folyóiratban kerültek közlésre.
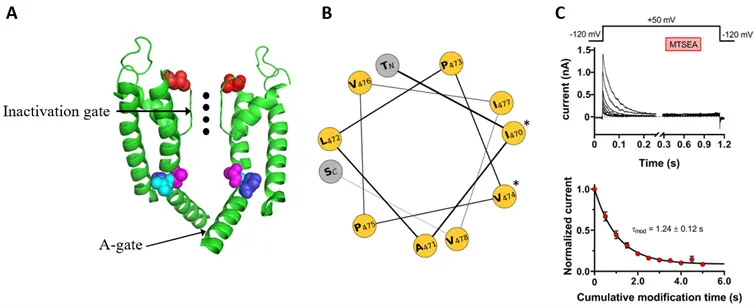
Ábra. (A) A Shaker feszültségfüggő káliumcsatornák pórus régiójának (S5 és S6 hélixek, valamint a hélixeket összekötő pórus formáló hurkok) szerkezete. A jobb áttekinthetőség érdekében az ábra csak két szemközti alegységet mutat szalag reprezentáció formájában. A színessel jelölt aminosav-oldalláncok: T449 (piros), A471 (lila), L472 (cián) és 473 (kék). A szelektivitási szűrőben lévő káliumionokat fekete pontok jelzik. (B) A pórus régió helikális vetülete. A spirális kerék jobb oldalán *-gal jelölt aminosav-oldalláncok az ioncsatorna nyitott állapotában a pórus felé néznek. (C) Az MTSEA gátolja a T449A/A471C Shaker ioncsatornákat inaktivált állapotban. (Felül) A feltüntetett feszültség-lépcső protokollal rögzített reprezentatív áramgörbék. Az MTSEA impulzus időzítését és időtartamát a satírozott sáv jelöli. (Alul) Az egyes depolarizáló impulzusok során mért csúcsáramokat az MTSEA alkalmazása előtti utolsó impulzus csúcsáramára normalizáltuk, átlagoltuk és ábrázoltuk a kumulatív módosítási idő függvényében. Az így kapott pontokra illesztett egyexponenciális függvény segítségével a gátlás sebessége meghatározható.